![Transcription factories in a Hela cell [from Cook PR (1999) Science 284, 1790]](../images/pombo.png)

('15nucbm'; Peter.Cook@path.ox.ac.uk; http://users.path.ox.ac.uk/~pcook)
Objectives
The material covered in this lecture, coupled
with the recommended reading, should enable you to:
• appreciate the size of nuclei and the different structures in it
• describe the different structures (types of chromatin, membrane, pores, nucleolus)
• understand how long DNA molecules are packed
into nuclei
• describe the crucial nuclear functions (replication,
transcription, repair, recombination)
Overview
Most conspicuous organelle in cell; diameter 2-10 micrometer (Fig);
arguably most important (contains genome).
Function: contain/protect genome, carry out replication, transcription,
repair, recombination, ribosome production.
Genes surrounded by envelope of 2 membranes.
Nuclear pores pierce envelope and allow communication between nucleus
and cytoplasm.
Generally little internal structure visible in LM,
apart from nucleoli (where rRNA synthesized, ribosomes
assembled).
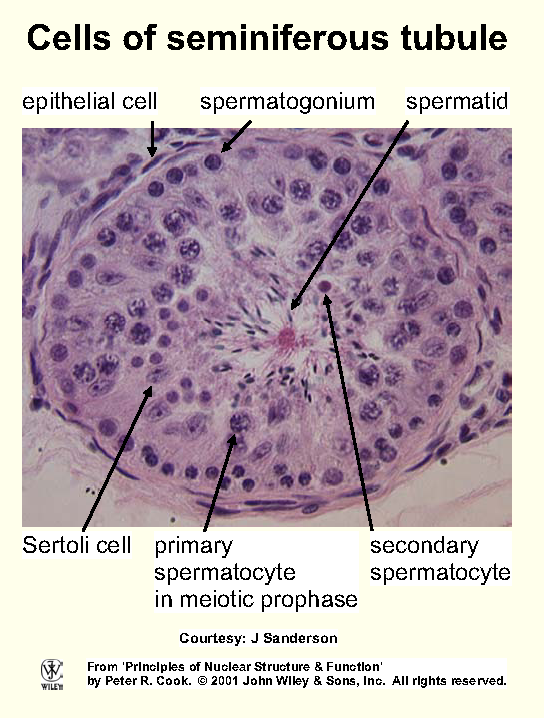
Most of rest filled with chromatin, recognized by reaction
with basic stains (eg methylene blue, hematoxylin). Cellular
components that bind basic and acidic dyes are termed basophilic and acidophilic.
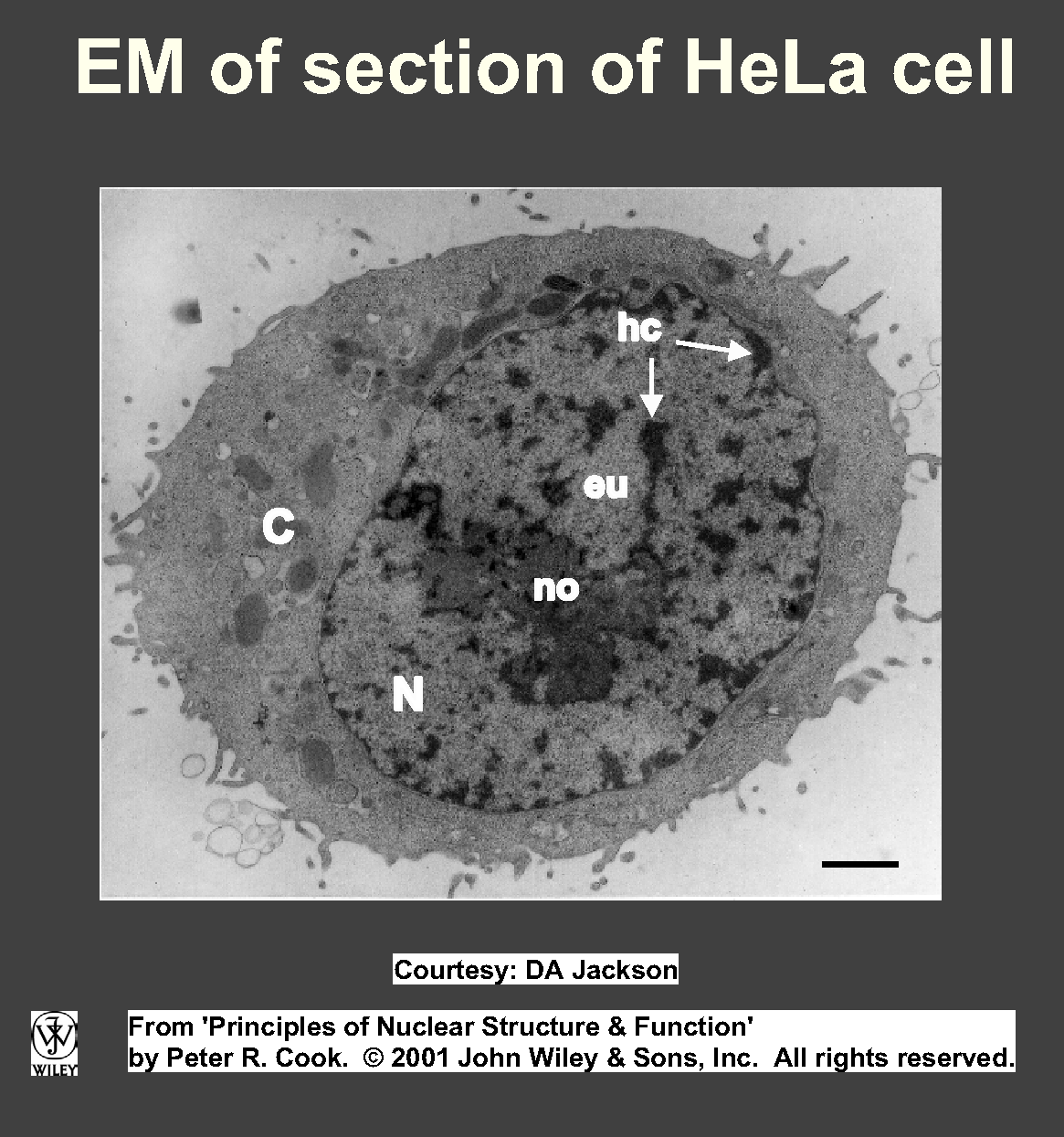
Chromatin commonly divided into 2 types: euchromatin (dispersed, occupies most nuclear volume), heterochromatin (densely
packed, structure more like that in mitotic chromosomes; Fig). Heterochromatin often condensed against envelope/nucleolus, can aggregate into densely-staining - often internal - chromocentres.
2 kinds of heterochromatin: constitutive (never expressed, often
contains short DNA repeats), facultative (expressed
in some cell lineages) -- example is X chromosome in female cells of mammals where one X (selected randomly early during development) is heterochromatic,
(almost entirely) transcriptionally inactive, forms dense Barr body
under nuclear membrane whilst the other (with essentially same DNA sequence) is euchromatic and inactive.
Size and structure
Typical human nucleus: ~10 µm diameter; occupies small fraction cell (6% vol in liver where 22% is
mitochondrial).
Nuclei containing different amounts DNA may have roughly same size, conversely, those with same amount DNA can have very different sizes
(eg nucleus in resting human lymphocyte is ~5 µm
dia, in migratory neuroblast >20 µm).
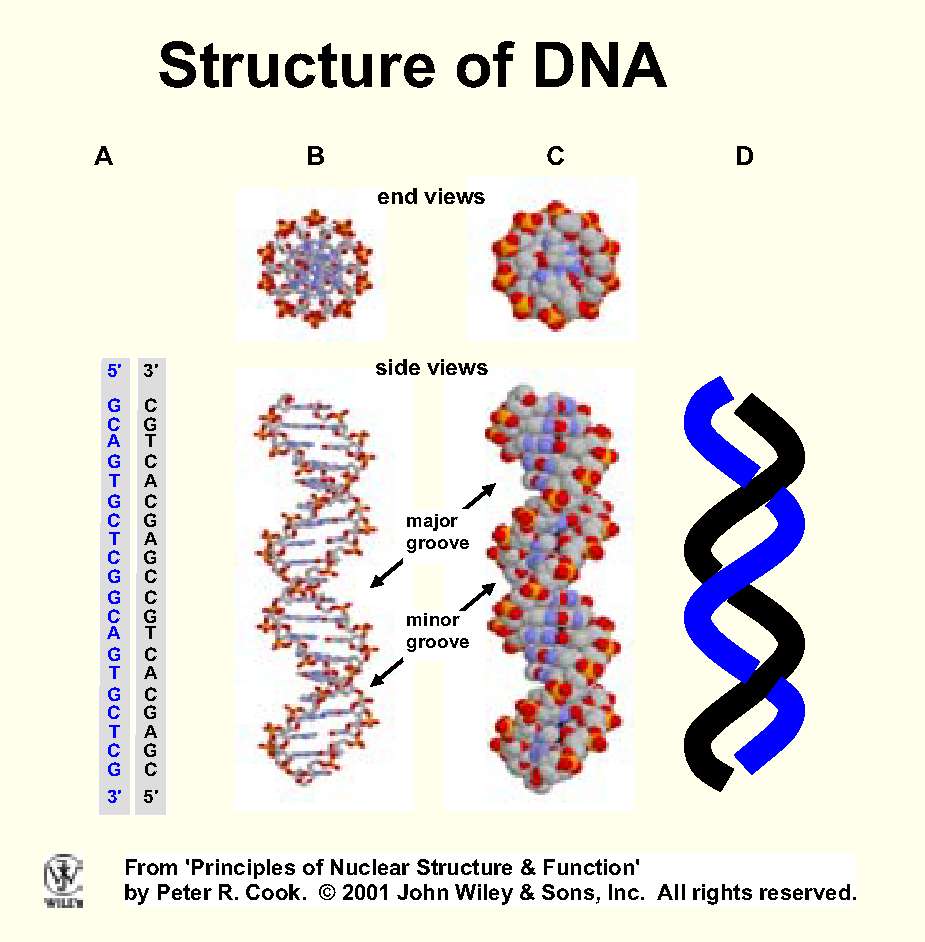
DNA in a human chromosome is arguably the longest and most important biomolecule (Fig)
Widely assumed there are ~3,000,000,000 bp (or 3,000 Mbp) in a haploid
human nucleus.
Typical human chromosome contains ~100 Mbp DNA; 2 nm wide, 3.4 cm long
when stretched out; 3,000 Mbp DNA stretches 2 m. [From
number of bp x distance between two in Watson-Crick structure (ie ~0.34
nm).]
Dimensions of molecules with such length-to-width ratios are far outside anything found in everyday world. Analogy:
kite string (~2 mm dia) is a millionfold wider than DNA fibre - if
had same length-to-width ratio as 100 Mbp of DNA, it would be 30 km long!
String equivalent to 6000 Mbp in diploid nucleus stretches 2000 km (London-Rome).
Packing problem - reduce contour length ~10,000x.
Easy to imagine packing controls access
polymerases to DNA.
DNA strong acid - neutralized - histones H1, H2A/B, 3/4 (contain positively charged amino acids - lys,
arg).
Histones highly conserved (2 differences in aa sequence of H4 from peas,
cows).
Non-histone chromatin proteins (many bind to specific DNA sequences).
DNA coiled around nucleosome, into solenoid (?), loops,
chromosome domains/territories (Fig).
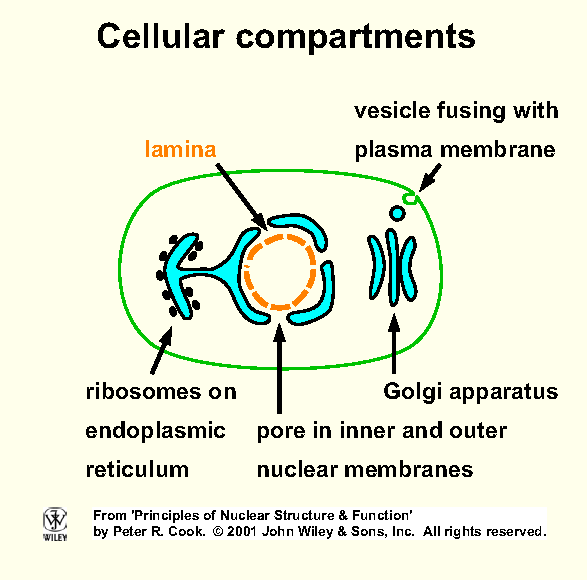
Nuclear envelope - distinguishes eu- from pro-karyotes (Fig).
Double membrane (inner, outer)
separated by 20-40 nm (perinuclear space). In EM, outer membrane
continuous with that of endoplasmic reticulum (ER); often outer surface
studded with ribosomes (rough ER). Space between nuclear
membranes continuous with lumen of ER.
A nuclear lamina - determines nuclear shape
- underlies nucleoplasmic side of inner membrane. Fibrous mesh of
lamin proteins, members of intermediate filament family (family contains
keratins, vimentin).
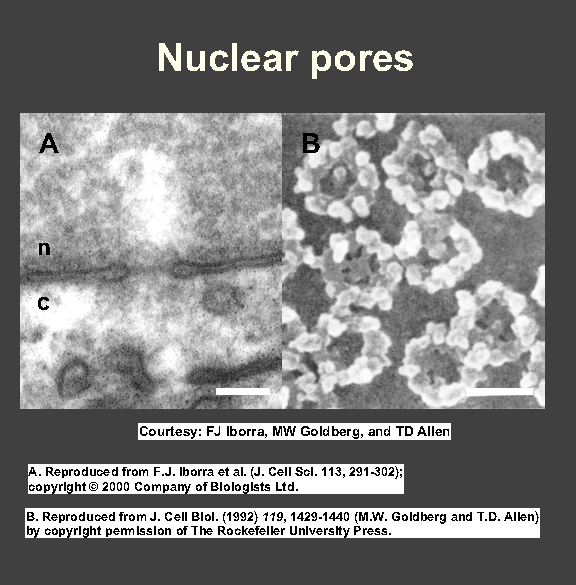
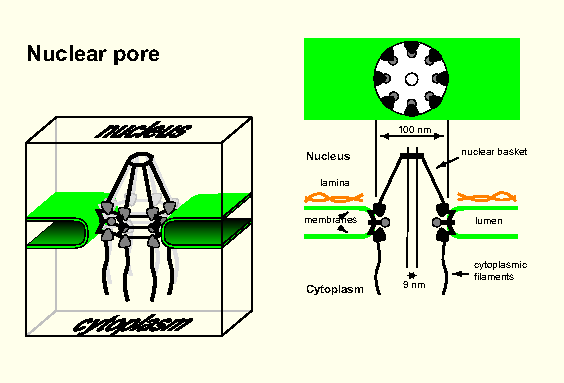
Nuclear pores - each surrounded by a nuclear pore complex - pierce membrane (Fig, Fig); gates allowing in/out.
Each pore enormous (~125 x 106 D; ~100 different polypeptides).
8 large protein granules arranged in a circle around hole (internal/external dia ~80/~120 nm); 8 fibrils attached to cytoplasmic
side of the ring, 'basket' on other side.
Central hole: aqueous ~9-nm channel allowing diffusion through membrane (proteins >9 nm cannot diffuse in/out).
Traffic density very high - in human cells ~50 histone molecules + ~100
ribosomal proteins enter every min through each pore, while ~2 ribosomal
subunits (dia ~15 nm) exit!
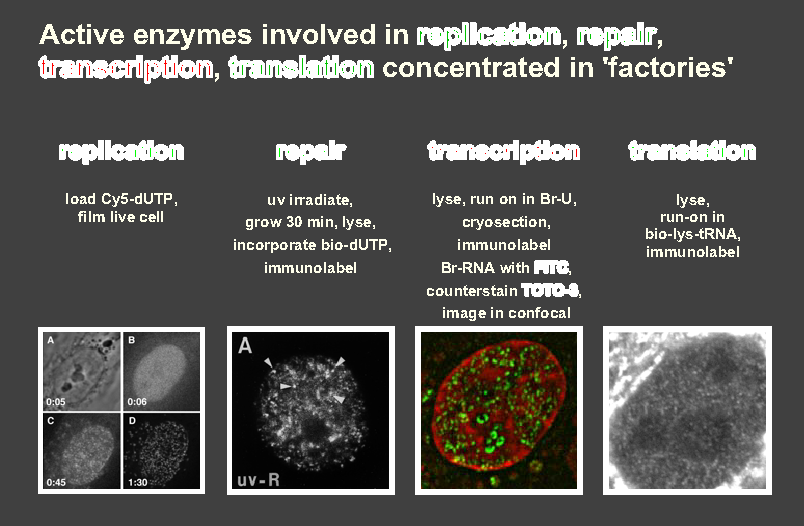
Functions - gene transcription, replication, repair, recombination.
Label newly-made RNA and DNA with appropriate precursors (eg uridine and
UTP/thymidine and TTP, tagged with 3H, 32P, Br, biotin)
to label nascent RNA/DNA. Note phosphorylated precursors cannot cross cell
membrane.
Active polymerases concentrated in discrete transcription/replication 'foci'
or 'factories' (Fig).
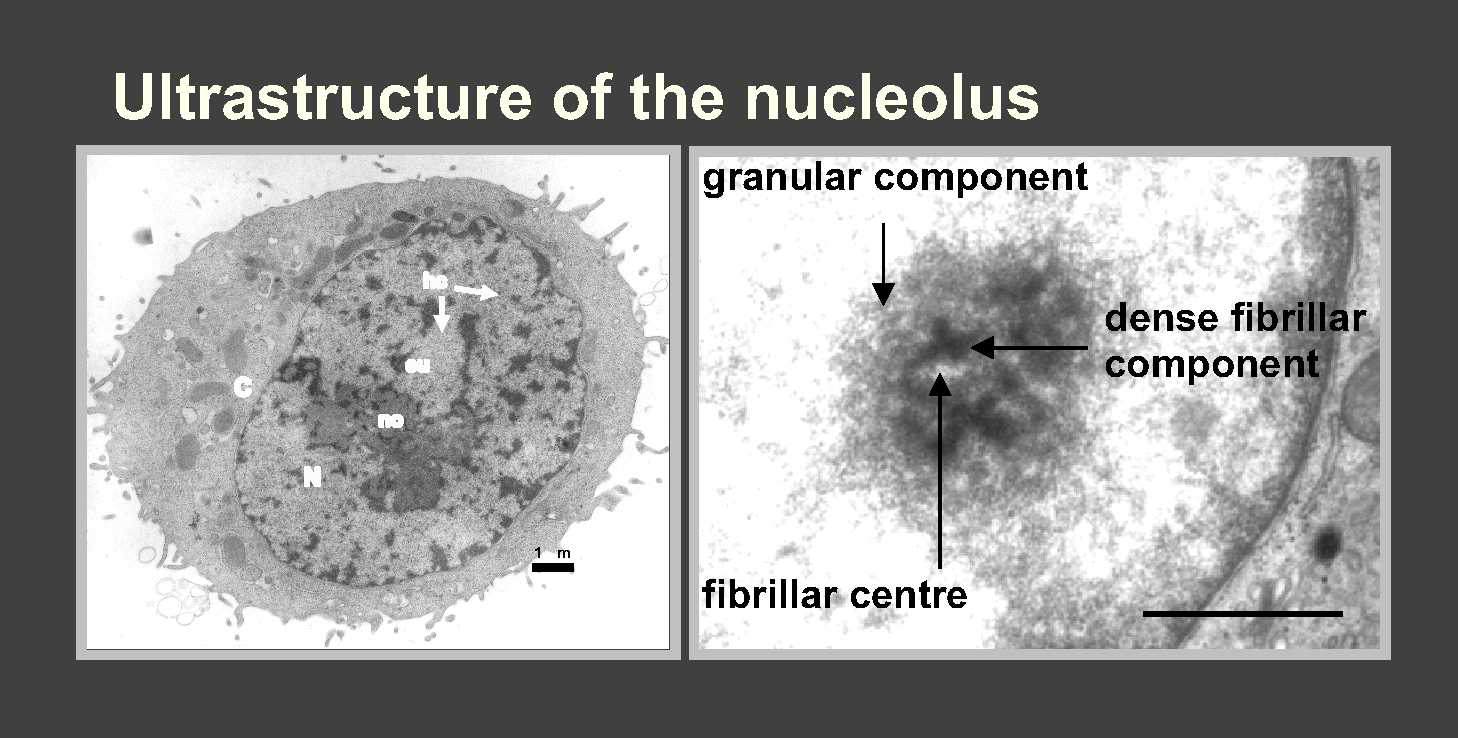
28, 18, 5S rRNA made in nucleolus, a ribosome-producing factory
(Fig).
The nucleolus is the most prominent cytological feature of nucleus, with
high concn RNA but little DNA. Its size reflects activity - small in dormant
cells, can swell to 25% nuclear vol in cells actively making protein.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Reference: Ch 4,5 Alberts, B. et al. (2014). 'Molecular
Biology of the Cell'. 6th Ed. Garland. [see also PubMed].
Pollard, T.D., Earnshaw, W.C., and Lippincott-Schwartz,J. (2007). 'Cell Biology'. 2nd Ed. WB Saunders/Elsevier.